
Meiosis
In most Sellaphora species, it is very easy to recognize pairs in meiotic prophase, because of the displacement of the chloroplasts (described on the previous page) and because the nucleus is larger and more conspicuous than at any other time during the life cycle, and, of course, because cells are paired.
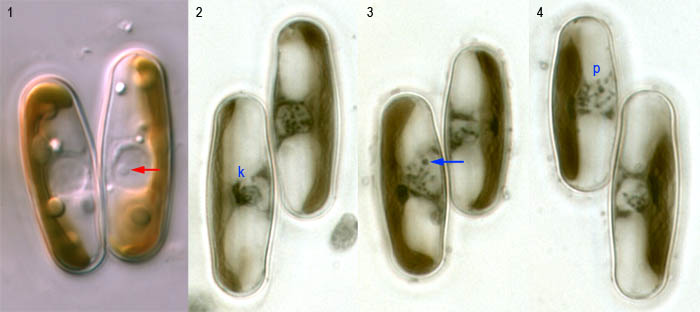
In living cells, the meiotic nucleus can be seen to be surrounded by a thick rim (Fig. 1), which probably consists of massed golgi bodies. During the early stages of meiosis, the nucleolus is prominent (Fig. 1, arrow) and can be seen to move around within the nucleus.
In early prophase, the chromosomes become visible within the nucleus and condense into a dense mass; the remainder of the nucleus becomes transparent and apparently watery. These features can sometimes be seen in living cells, but are more obvious in stained preparations (Figs 2–4: these were made by fixing sexualized populations with Flemming's solution and staining with haematoxylin). Fig. 2 shows a 'synizetic knot' of chromatin within one nucleus (left), separated from the nuclear envelope by an empty space. A less dense knot is present in the left-hand cell in Fig. 3 and here a nucleolus can be seen (arrow) at or near the tip of a chomosome. These cells are in zygotene, when homologous chromosomes pair. Later, in pachytene (Fig. 4, p), the chromosomes shorten and thicken and begin to disperse out of the synizetic knot. We have not yet published a detailed account of meiosis in Sellaphora, but the pictures shown above may be compared with meiosis in Navicula (Mann & Stickle 1989) and Placoneis (Mann & Stickle 1995), or the older very careful account of Cymbella by Geitler (1927).
Unfortunately, it has not yet been possible to count the chromosomes in any Sellaphora species. This is not a special problem of Sellaphora: it is a consequence of two factors (1) the structure of metaphase (mitoitc or meiotic), in which chromosomes cluster densely around a narrowly cylindrical spindle and cannot be counted separately; and (2) the silica cell wall prevents use of normal squash methods. Figures 2–4 suggest that the chromosome number is not small (n is probably >10) in S. blackfordensis.